I have seen the future and it is the Midbrain Reticular Nucleus
November 14, 2019
In lab meeting this week, we read a biorXiv paper by Nick Steinmetz, Peter Zatka-Haas and colleagues.
Approach: The authors used Neuropixels probes to measure the responses of 30K neurons in mice reporting spatial judgments about visual stimuli. The very large dataset was accompanied by careful reconstructions of electrode tracks (using a combination of histological and electrophysiological data) that allowed them to localize the neurons to 42 brain regions. There’s a lot there! It was a real tour-de-force and the lab had to think hard to define 3 main take-home messages:
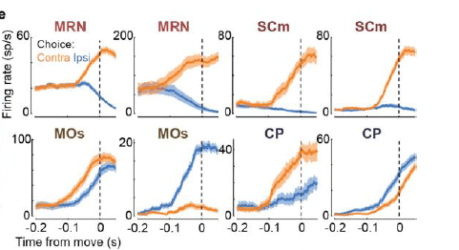
1. Choice signals (more on what that means later) were sparse and widely distributed. Interestingly, they were plentiful in deep structures including the midbrain reticular nucleus (MRN, see example below). The MRN is not really known as being a hot spot for decision-making, highlighting an advantage of using probes that allow multi-area recording that extends to subcortical structures. I predict more studies will measure activity there in the future (note the appealing bidirectional modulation in the examples below).
2. Movements explained the majority of trial-to-trial variance brain-wide (yep).
3. The effects of arousal were interesting. Heightened arousal tended to suppress activity in the pre-trial period for cortical neurons, and enhance activity in the pre-trial period for deeper structures neurons. Moreover, using this pre-trial activity to generate an “engagement index” allows one to predict pretty well whether the mice would “go” or not much later in the trial.
Skeptics corner: Overall, this was a great paper; the broad survey of brain areas led to some surprising observations. Moreover, because the design here was to explore the brain broadly on an interesting task, the authors weren’t aiming to support or refute a specific hypothesis about mechanisms supporting the task. Still, there were some observations that we puzzled over.
- The responsiveness to visual stimuli in primary visual cortex was low. This was not totally unexpected and has been reported in other studies. Still, in figure 2d, only about 18% of V1 neurons were responsive during contralateral correct trials and a lot of the response was action driven (as the authors acknowledge). This is really low. Admittedly, we weren’t sure what to expect: stimuli were perhaps out of the receptive field of many measured neurons, and weren’t optimized for the spatiotemporal frequency nor orientation preferences for each cell. We also weren’t sure how to compare the metrics of visual responsiveness in Fig. 2 with the one in Fig. 3. In a linear encoding analysis in which neural activity was fit with vs. without a “vision kernel”, many more neurons were classified as “visual encoding” (nearly 30% in Fig. 3d). I liked their encoding analysis and wasn’t sure why it would uncover visual encoding that was missed in a more basic analysis, especially because the latter included neurons that were responsive any time in the trial so would seem to be a looser criteria.
- The paper made me ponder what we should consider to be a “choice” signal in the brain. The authors did something I consider fair, which is to separate “action” signals, which don’t distinguish left vs. right responses, from “choice” responses, which do distinguish the two. This meant that the total number of “choice” neurons was really low: they were most numerous in the MRN (!) and the zona incerta, but even there barely exceeded 5%. This fits with other observations, including our own, about the paucity of “choice” signals. I think that the authors are actually being fairly generous with what they consider a “choice” neuron (moreso than we were, in fact). If I understood the analysis correctly, the “choice” neurons are the ones that respond differently to left vs. right movements of the wheel. So many of these might actually be neurons that are involved in the preparation and execution of the complex set of limb movements needed to report the choice. Should there be another way of defining “choice” neurons? Do we expect to see some neurons in the brain that reflect the choice independent of the action used to report it? If we wanted to find such neurons, what would be the right experimental design? I think such neurons should exist. For instance, I might decide I want a coffee but not plan on whether I will walk or bike to the cafe until much later.
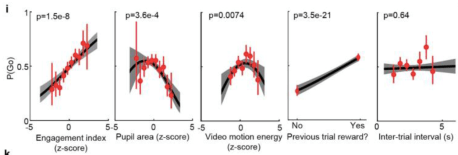
A final thought: there is a lot of really nice stuff in the supplemental and extended data. I recommend Extended Data Figure 9i (below). This demonstrates that in fact there are lots of ways to predict whether the animal will “go” or not late in the trial based on what it did early in the trial (or even on the previous trial). Kinda reminds me of this poster.